
Ceres is the Roman Goddess of agriculture, crops and Earth’s fertility; Ceres is also a dwarf planet between Mars and Jupiter, whose existence was predicted by proxy long before its actual observation and discovery.
CERES, an ERC (now UKRI) Advanced Research Grant aims to develop new biochemical and isotopic understanding of modern, past and future microbial processes in peatland, among the most important stores of organic carbon on Earth. In doing so, it will explore three major, inter-related questions about life and our planet.
How and why organic matter is made and how that governs its fate.
How Earth’s climate, environment and life co-evolved, especially during times of rapid change.
How systems respond to perturbations and how that response is dependent on the rate of change.
These disparate questions are held together by the central role of organic matter in life, the environment, energy and the Earth. It is what all life is made of, and its composition can be used to reconstruct the history of life. Its production through photosynthesis produces oxygen, its burial removes carbon dioxide, and its degradation governs the fate of numerous elements on the Earth’s surface, from sulfur and nitrogen to iron, mercury and arsenic.
Despite the study of organic matter in the environment being an over 50-year old discipline, peatlands remain understudied, and those studies have focused on temperate and polar peatlands, with tropical peatlands being long neglected. Consequently, their investigation will not only be exciting and novel, with initial work already providing tantalizing insights into unique microorganisms and biogeochemistry, but will be a platform for understanding the formation, diversity and persistence of organic matter, with a focus on the bacteria and archaea that govern either its preservation or its degradation into the greenhouse gases carbon dioxide and methane. Furthermore, by focusing on sites that have experienced disruption – recently, on millenial timesclaes and through Earth history – we will gain new insights on how systems respond to change across multiple timescales. When is balance maintained and when is it catastrophically broken? And when broken, what are the consequences of that period of chemical and biological disequilibrium and how long does it take for balance to be restored.

**************************************************************************************************
CERES is about the past and the future. It is about exploring new environmental settings. It is about new methods to interrogate the adaptations of microorganisms to their environment. And it is about new isotope approaches to unlock past and future microbial metabolism. We will build a diverse, inclusive and international team to collaboratively co-create a research plan that explores all of these aspects of organic geochemistry and Earth systems.
We will integrate analytical innovation, biogeochemistry in modern contexts, and geological archives to holistically evaluate how climate change affects peatland carbon cycling across multiple timescales. Greenhouse gases shaped Earth history, impacting both climate and ecosystems; ongoing anthropogenic emissions are doing the same. These include impacts on the microbially mediated processes that govern the chemical state of our planet and act as climate feedbacks. Soil microorganisms, for example, account for the largest natural methane flux, and yet these processes remain poorly understood, mediated by multiple environmental factors. Insight can be derived from geological archives that document the timescale-dependent responses of biogeochemical systems to environmental perturbations. Such studies on peat and lignites provide tantalising insights into climate-driven disruption of the carbon cycle, but the underlying mechanisms remain unresolved. This critical knowledge gap arises from our inability to determine the isotopic signatures of the most diagnostic biomarkers. Therefore, we will:
1) Develop new instrumentation for the isotopic determination of large bacterial and archaeal biomolecules. This is a transformative expansion of our biomarker toolkit as our analytical window expands from low to highly diagnostic compounds.
2) Learn from and share with international colleagues new approaches in lipidomics to understand the nature of archaeal and bacterial adaptation.
3) Refine existing methods in peatland organic matter composition, adapting them for the investigation of both highly degraded peats and tropical peats.
3) Apply these new methods to modern peatlands, examining biomarker isotopic compositions in the field as well as in experiments in response to manipulation of pH, temperature and substrate.
4) Apply our new biomarker methods and understanding to the geological past. Working across decadal to multi-million year timescales, we will unlock the mechanistic controls underlying ancient reorganisations of peatland carbon cycling. From dramatic climate change events of the past to monsoon-driven drying of Holocene wetlands to the catastrophic destruction of peatlands through exploitation of our landscape, we will explore the signature of catastrophic change on these ecosystems, as written in their molecular compositions.
Through these WPs, CERES will probe how microbial metabolism, and hence biogeochemical cycles, operate(d) on the Earth today, through its history, and in response to rapid global warming.
**************************************************************************************************
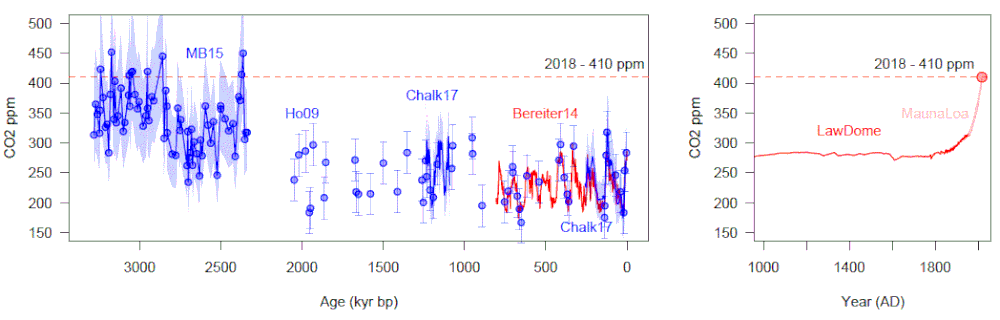
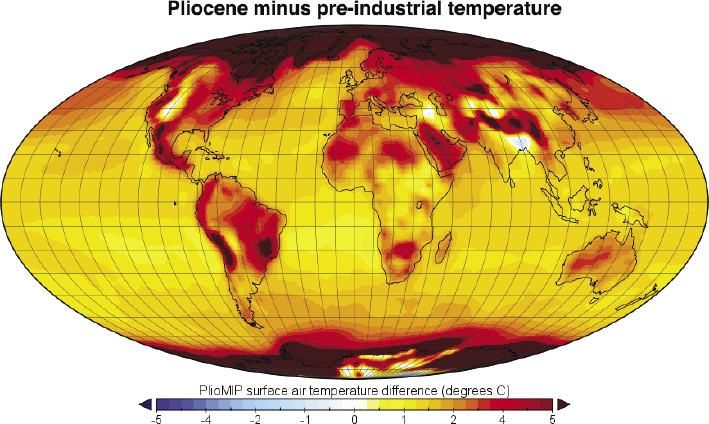
Overview from the project proposal: Microbially mediated biogeochemical processes in terrestrial settings are critical to governing greenhouse gas emissions. The modern carbon soil reservoir exceeds that of terrestrial vegetation and the atmosphere combined (Crowther et al., 2019), and soil microorganisms annually cycle 1/3 of the carbon photosynthesised and account for the largest natural methane flux. In doing so, they govern the chemical and climatic state of our planet. And yet these processes remain poorly understood, mediated by a range of environmental factors that respond to climate change at different rates (Liu et al., 2020). Such responses are crucial to dictating whether terrestrial systems act as positive or negative feedbacks to climate change and whether a system is vulnerable to tipping points (i.e. Steffen et al., 2018). We have explored the long-term biogeochemical response of these systems to climate change using ancient peatland deposits. Initial results, however, are inconclusive because we could only crudely reconstruct ancient biogeochemistry. This project will transform our capacity to probe such responses – at decadal to centennial scales in Holocene peat; and at millennial to million-year timescales in Cenozoic lignites. We will develop transformative new instrumentation for the isotopic analysis of heretofore inaccessible biomolecules and use this to measure the stable hydrogen and carbon isotopic composition, and by extension the metabolism, of microbes in modern and ancient environments. In doing so, we will determine the bacterial and archaeal response to climate change and the associated impact on biogeochemical cycles.


















{kind=link}